


Sélection inverse, un accélérateur de création variétale
-
 Les NBT recouvrent de nombreuses techniques différentes et ne visent pas toutes à influencer durablement l’expression des gènes des plantes. La sélection inverse n’est ainsi qu’une étape transitoire dans la constitution de nouvelles variétés, et s’appuie sur d’autres new breeding techniques.
Les NBT recouvrent de nombreuses techniques différentes et ne visent pas toutes à influencer durablement l’expression des gènes des plantes. La sélection inverse n’est ainsi qu’une étape transitoire dans la constitution de nouvelles variétés, et s’appuie sur d’autres new breeding techniques.
L’intérêt premier de la sélection inverse est de pouvoir obtenir rapidement des semences hybrides F1.
Une semence F1 est une lignée hybride de première génération, c’est-à-dire qu’elle est issue de deux lignées parentes homozygotes (les allèles sont tous les mêmes) ayant chacune leurs qualités. L’hybride F1 est donc hétérozygote (des allèles différents), profitant à la fois des avantages sélectifs de ses deux parents et de la vigueur hybride (l’hétérosis), qui rendent l’hybride particulièrement performant pour la culture. Cependant, la génération suivante (F2) ne conservera pas le génotype cumulant tous les avantages de la F1. La reproduction sexuée entre plants hétérozygotes crée des descendants ayant un patrimoine génétique différent des parents : ils n’auront donc plus l’ensemble des caractéristiques intéressantes.
Il faut donc recroiser des lignées pures pour produire de nouveaux hybrides F1. Or, ces lignées pures sont elles-mêmes issues d’un long travail de sélection conventionnelle. La sélection inverse permet d’accélérer ce processus.
La sélection inverse, un procédé qui fait appel à d’autres NBT
La sélection inverse est constituée de plusieurs étapes :
1. Blocage de la méiose : chez un hybride F1 que l’on veut reproduire, la recombinaison méiotique va être inhibée, grâce à l’inactivation du gène qui contrôle ce processus, par exemple à l’aide d’ARN interférents, de CRISPR/Cas9, TALENs, etc. Le but est d’éviter le brassage intrachromosomique au moment de la production de gamètes. Chaque gamète est donc constitué de chromatides identiques à l’une des deux lignées parentales de l’hybride F1.
2. Doublement du patrimoine génétique : les gamètes sont mis en culture pour obtenir des plantes haploïdes, qui n’ont qu’une seule version de chaque chromosome. Certaines plantes sont capables de doubler spontanément leur nombre de chromosomes, en créant une copie des premiers. Pour d’autres, il faut recourir à la colchicine, une substance capable de provoquer ce doublement des chromosomes et extraite des colchiques.
Les plantes obtenues, diploïdes (deux exemplaires de chaque chromosome) sont donc homozygotes (un seul allèle de chaque gène).
3. Reconstitution de l’hybride : les plantes diploïdes obtenues à l’étape précédente constituent de nouvelles lignées pures qui sont utilisées comme parentes d’une nouvelle génération F1. Il suffit d’identifier (notamment grâce à des marqueurs génétiques) deux lignées parentes qui, à elles deux, cumulent tous les gènes d’intérêt de l’hybride initial.
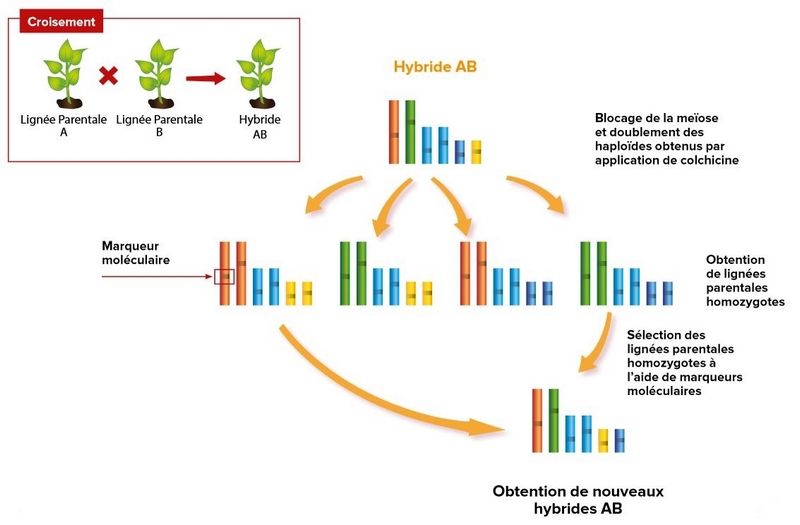
Schéma récapitulatif de la sélection inverse (Horizon biotechs)
Grâce à cette sélection inverse, en partant de « l’enfant » pour reconstituer le patrimoine génétique des « parents », il est possible de générer rapidement de nouveaux hybrides F1 équivalents aux originaux. Facilitant notamment la sélection et le maintien des patrimoines génétiques pour les obtenteurs, la sélection inverse peut s’avérer particulièrement utile pour les PME, aux moyens plus limités.
La sélection inverse est de fait adaptée à de nombreuses espèces de plantes consommées par l’homme : riz, maïs, tomate, brocoli, oignon, concombre… Toutefois, une limite demeure pour l’instant, liée à la complexité des génomes. Plus le génome est simple (celui du riz ne compte ainsi que 12 chromosomes et moins de 40 000 gènes), plus cette NBT est efficace. Pour les génomes très complexes, comme celui du blé, qui est hexaploïde (6 versions de chaque chromosome), cette technique est encore en cours de perfectionnement.

